Reading...
![]()
Play button
![]()
Play button
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
15 Cards in this Set
- Front
- Back
|
α-oxidación
|
consiste en la eliminación secuencial el carbono carboxílico en forma de CO2, por lo que el Cα del Ácido graso de partida se convertiría en el carbono carboxílico del siguiente ciclo
|
|
|
ω-oxidación
|
esta se da en los peroxisomas. Esta oxidación va a actuar a nivel del último carbono de la cadena. En este caso la oxidación no va a suponer liberación de carbono sino que es oxidación y conversión, de ese extremo carbono a carboxilo. Suponen la hidroxilación del extremo opuesto al carboxilo de un ácido graso, cuya oxidación posterior generaría un nuevo extremo carboxílico, con ello se obtiene un di-ácido.
|
|
|
β-oxidación
|
esta es la que van a realizar las mitocondrias, se trata de producir una oxidación entre los carbonos 2 y 3, es decir, entre los Cα y Cβ. Cuando esa oxidación se produce va a producir una ruptura a nivel de ese enlace y el Cb se va a convertir en el C1 del ácido graso acortado. Por lo que consiste en la eliminación secuencial de fragmentos bicarbonatados desde el extremo carboxilo (el enlace entre Cα y Cβ). La idea fundamental sería que al final de cada ciclo de β- oxidación, el Cβ del ácido graso de partida se convertiría en el “Carboxilo” del siguiente ciclo.
|
|
|
ACTIVACIÓN ENERGÉTICA DE LOS ÁCIDOS GRASOS
|
Los ácidos grasos precisan ser activados energéticamente para poder sufrir la β-oxidación, el enzima responsable es la Acil CoA grasoSintetasa que gasta el equivalente a 2 ATP para convertir el ácido graso en Acil CoA graso.
Es un proceso en el cual partimos de un ATP, que transfiere una adenosina a la cadena del carboxilo, por lo que tenemos una amp acil graso, luego se libera un pirofosfato el cual mediante pirofosfatasa inosrgánica se convierte en dos fosfatos inorgánicos. Cada vez que este proceso ocurre supone el gasto aproximado de 2 ATP. |
|
|
Una vez tenemos el Acil CoA graso este tiene que atravesar dos membranas, ya que la β-oxidación se produce en el interior de las mitocondrias. Existen 2 casos:
|
- Ác. Grasos con menos de 12 C: que atraviesan la membrana interna mitocondrial por un mecanismo que no requiere carnitina, mediante un sistema de difusión básica.
- Ác. Grasos con más de 12 C: necesitan un sistema para atravesar la membrana que requiere diversos enzimas y un transportador. Este tipo de ácidos grasos requieren un tipo de transporte Carnitina-dependiente, que requiere dos actividades enzimáticas. |
|
|
El transporte Carnitina-dependiente consiste en lo siguiente:
Primero el ácido graso es activado en la membrana externa por la Acil-CoA Sintetasa, después en el espacio intermembrana la Carnitina Aciltransferasa I o CAT I ó CPT I, transfiere el acilo desde el CoA a la Carnitina, dando así Acil-Carnitina. Posteriormente la Acil-Carnitina se transporta a la matriz en un antiporter con Carnitina, situado a nivel de la membrana interna mitocondrial, que mete hacia el interior, hacia la matriz mitocondrial Acil-Carnitina al mismo tiempo que saca hacia el exterior Carnitina. Y una vez dentro de la matriz mitocondrial tenemos al segundo enzima la Carnitina Aciltransferasa II o CAT II o CPT II la cual transfiere el acilo desde la Carnitina al CoA, obteniéndose así Acil-CoA en la matriz mitocondrial. Por lo que el CAT II revierte la acción del Cat I. |
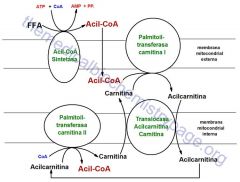
|
|
|
REACCIONES DE LA β-OXIDACIÓN .Cada ciclo de β-oxidación implica 4 reacciones sucesivas:
|
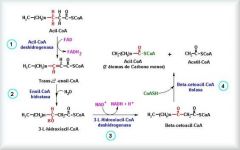
|
|
|
RENDIMIENTO DE LA β-OXIDACIÓN: Ác. Grasos saturados de cadena PAR
|
En un ácido graso de cadena par con n Carbonos el número total de ciclos de β-oxidación necesarios para su oxidación completa sería: (n/2 -1). Cada ciclo producirá: 1 FADH2 + 1 NADH + 1 Acetil-CoA, excepto el último ciclo que produciría 2 Acetil-CoA.Nº ATP= (n/2-1) X 5 ATP + (n/2) X 12 ATP – 2 ATP
|
|
|
RENDIMIENTO DE LA β-OXIDACIÓN: Ác. Grasos saturados de cadena IMPAR
|
Los ciclos de β-oxidación producen el mismo rendimiento que los saturados de cadena par excepto en el último ciclo que produce: 1 Propionil-CoA + 1 Acetil-Coa, en lugar de 2Acetil-CoA. Dicho Propionil-CoA se transforma en Succinil-coA ingresando hacia la mitad del ciclo de Krebs y rindiendo solo 6 ATP. La ruta que sigue hasta conseguir el Succinil-CoA implica tres pasos:
|
|
|
Propionil-CoA se transforma en Succinil-coA ingresando hacia la mitad del ciclo de Krebs y rindiendo solo 6 ATP. La ruta que se sigue hasta conseguir el Succinil-CoA implica tres pasos importantes
|
1- Carboxilación: el enzima Propionil.CoA Carboxilasa, usando HCO3- como fuente de carbonos y gastando 1 ATP, carboxila el carbono α del Propionil-coa danado D-Metilmalonil-CoA.
2- Racemización: una epimerasa ó racemasa cambia la posición del CoA convirtirendo el D-metilmalonil-CoA en L-Metilmalonil-CoA. 3- Mutasa: el enzima Metilmalonil-CoA mutasa, que requiere vitamina B12, transfiere el Carboxil-CoA desde el carbono α al metilo, convirtiendo el D-Me |
|
|
en la oxidación completa de un ácido graso lineal saturado de cadena impar con “n” Carbonos:
|
Nº ATP = (n-3/2) X 5ATP + (n-3/2) X 12ATP + 3ATP.
|
|
|
Grasos IN-saturados
|
Todos los dobles enlaces (insaturaciones), de los ácidos grasos naturales presentan conformación CIS. La conformación CIS de estos enlaces no es sustrato para los encimas de la β-oxidación, requiriéndose enzimas especiales para poder continuar la oxidación de los ácidos grasos una vez alcanzamos las insaturaciones.
En principio, los ácidos grasos insaturados sufrirían ciclos normales de β-oxidación, hasta encontrarse con el docle enlace en CIS. Durante el ciclo de β-oxidación la “maquinaria” se encuentra que hay un doble enlace que puede estar en una posición de un carbono impar o en una posición de un carbono par. |
|
|
Ácidos grasos Cis-Δ9 o Cis-Δimpar:
|
- Tras tres ciclos completo de β-oxidación, tendríamos un Cis-Δ3.
- El enzima Enoil CoA Isomerasa isomerizaría el Cis-Δ3 a Trans Δ2, el cual es un sustrato para el segundo enzima de la β-oxidación (Enoil CoA hidratasa), completándose este ciclo, aunque este sería incompleto al haberse saltado el primer paso del ciclo, por lo que no produce FADH2. - el resto de la cadena sufriría 4 ciclos completos de β-oxidación más hasta el final. |
|
|
Ácidos grasos Cis-Δ12 o Cis-Δpar:
|
- Tras tres ciclos completos de β-oxidación, tendríamos un Cis-Δ3, el enzima Enoil CoA Isomerasa isomerizaría el Cis-Δ3 a Trans Δ2, completándose este ciclo, aunque sería incompleto puesto k no produce FADH2. Hasta este punto seria idéntico al del Oleoil CoA
- Tras producirse la primera reacción del siguiente ciclo, en la que la Acil-CoA deshidrogenasa produciría 1FADH2, tendríamos un Trans-Δ2, Cis-Δ4. - El enzima 2,4 Dienoil-CoA Reductasa gastando 1 NADPH convierte los enlaces Trans-Δ2, Cis-Δ4 en uno solo Trans-Δ3 - La Enoil-CoA Isomerasa isomeriza el enlace Trans-Δ3 a Trans-Δ2, completándose este ciclo. Este si sería un ciclo completo ya que se inició con la primera reacción de la β-oxidación produciendo FADH2 - Finalmete se realizarán 3 ciclos completos de β-oxidación hasta el final |
|
|
Rendimiento
|
- Por cada enlace Δimpar que haya, supondría la ausencia de producción de 1 FADH2, en ese ciclo.
- Los enlaces Δpar no afectan al rendimiento del ciclo así que quedarían ciclos completos Según lo anterior el numero completo de ciclos sería: (n/2-(1+nº Δimpar)) Por lo tanto, el ATP sería: (n/2-(1+nº Δimpar)) x 5ATP - En todos los casos obtendríamos n/2 moléculas de Acetil-CoA. Cada molécula rendiría por Krebs 12 ATP: (n/2) x 12 ATP - Habría que restar los 2ATP de la activación energétia de de Acil-CoA. - Por lo que la fórmula quedaría de la siguiente manera: Nº ATP= (n/2-(1+nº Δimpar)) x 5ATP + nº Δimpar x 3ATP + (n/2) x 12 ATP -2 ATP |